Comparison between Sarcophilus and Gulo, part 2
(writing in progress)
Although much has been written about the bite-force of Sarcophilus, and its role as bone-crunching scavenger, there is an obvious aspect to the dentition of this marsupial that seems to have been underplayed. And this is pertinent to our current comparison with Gulo.
The overlooked/underplayed aspect is that there seems to be nothing specialised about this dentition as such, w.r.t. bone-crunching. This stands in contrast to the dentition of e.g. Crocuta, which certainly is specialised for bone-crunching.
Sarcophilus achieves its formidable status as a bone-cruncher in three ways, which I have not seen stated clearly before:
- Large head and teeth relative to body size, which naturally boosts bite-force
- Extremely wide gape, ensuring that bones can be placed between the posterior-most molars, thus using leverage in a similarly crude way
- Short lifespan, which means that wear (breakage) of dental cusps matters relatively little because the dentition is not designed to last long.
At the risk of caricaturising Sarcophilus, let me put it this way:
Sarcophilus has remained unspecialised, because just as there has never been a niche for a large scavenger in Australia there has also never been a niche for a scavenger committed to a diet of bones. Essentially, Sarcophilus is just a scaled-up dasyurid with a particularly scaled-up head, but few qualitative modifications to the typical dasyurid design. By retaining the typical dasyurid pattern of short lifespan, it has got around the need to make its teeth durable in crushing bones. It instead just uses a crude tactic, biting hard on bones with teeth that are not particularly designed for such application, and accepting the inevitable damage to the teeth.
And by retaining a relatively small body and thus appetite, Sarcophilus is adapted to a continent with sparse and unreliable food owing to its nutrient-poor soils. A combination of small body (and thus limited food-demands) with large head (and thus versatility in taking large items as well as smaller ones) makes sense in the Australian context.
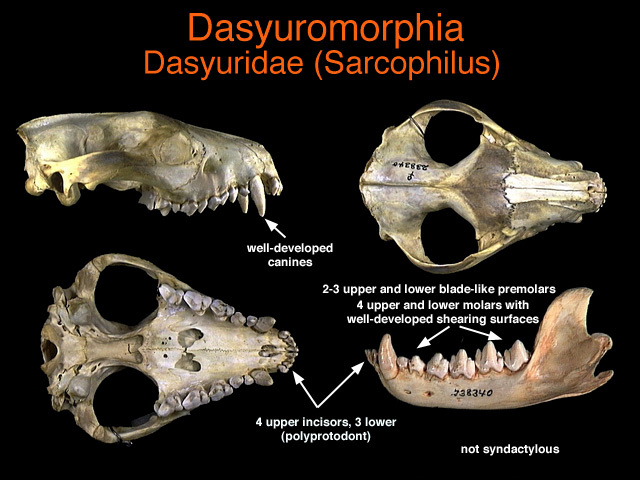
You will see from the photos below that Sarcophilus possesses neither the flat molars seen in Canis and Ursus, nor the conical premolars seen in Crocuta. Not only are its molars unremarkable, but they even retain the sharp cusps of a typically insectivorous mammal, scaled up to a size where they are far too big to be suited to insectivory but are instead used in a crude way on large items including hard ones. Sarcophilus does not even have a particular carnassial dentition along the lines of felids or Canis; instead its whole cheek-tooth row is carnassial but in a relatively imprecise and thus poorly-specialised way. It is consistent with this ‘crude’ approach to dentition that Sarcophilus does not bother to have a deciduous dentition while growing to maturity; it just crudely subjects its part-grown adult teeth to wear from the start, with little regard to keeping the teeth intact for as long as possible as most eutherians do.
In this way, Sarcophilus is functionally hyena-like ‘by default’, having adapted not its teeth as such but rather the crude parameters of the rest of the body and the pace of life.
By contrast, bone-crushers such as Crocuta have proportionately small heads and teeth, but use a far more ‘technically sophisticated’ solution to applying force, using ‘precision-tooling’ rather than brute force. In accordance with this, Crocuta has a great lifespan, the opposite of what is seen in Sarcophilus. And the simplicity of the approach of Sarcophilus, compared with the ‘artfulness’ of the approach of Crocuta, correlates with the great difference in relative brain size between these mammals.
The following several photos show how large the skull of Sarcophilus is relative to body size. The disproportionate size of the cheek-teeth is the main reason for its bite-force.
https://upload.wikimedia.org/wikipedia/commons/thumb/d/d4/Tasmanian_devil_skeleton.jpg/220px-Tasmanian_devil_skeleton.jpg
https://s-media-cache-ak0.pinimg.com/originals/e5/71/b8/e571b8e2f005a4dce30ca33dce84317e.jpg
https://www.wired.com/images_blogs/wiredscience/2009/05/3-tasmanian-devil-fossil_rm.jpg
https://upload.wikimedia.org/wikipedia/commons/5/55/Tasmanian_Devil_skeleton.jpg
http://www.nhc.ed.ac.uk/images/collections/mammals/marsupials/devilskel.jpg
The following four photos show that all of the cheek-teeth have cusps that are initially sharp in youth, before being broken by wear.
http://animaldiversity.org/collections/contributors/anatomical_images/family_pages/dasyuromorphia/dasyuridae/medium.jpg
https://upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Animaldentition_sarcophilusharrisii.png/220px-Animaldentition_sarcophilusharrisii.png
https://i0.wp.com/upload.wikimedia.org/wikipedia/commons/6/64/Cambridge_Natural_History_Mammalia_Fig_076.png
https://c1.staticflickr.com/6/5086/5316470850_d387a102a0_b.jpg
The following many photos show the whole skull. The great size of the canines certainly does show impressive specialisation for carnivory, but has little bearing on bone-scavenging. Look in particular for any sign of ‘crowding’ of the lower incisors, which as we’ll see in part 2 is a special feature of Gulo.
https://boneclones.com/images/store-product/product-608-main-original-1454455009.jpg
http://www.skullsunlimited.com/userfiles/image/variants_large_4392.jpg
http://lrrpublic.cli.det.nsw.edu.au/lrrsecure/sites/web/gondwana/Animal_Fossils_of_Gondwana/lo/fossils_06/graphics/skull4_wh_sm.jpg
http://img01.deviantart.net/68da/i/2015/239/a/5/tasmanian_devil_skull_by_lizardman22-d94j8wd.jpg
http://tasmaniantimes.com/images/uploads/skull1mooney.jpg
https://www.scammellauctions.com.au/catalogue/shop_image/product/595/thumb-13ae4099fd5e4af8bdefee83e0a08c4f.jpg
https://s-media-cache-ak0.pinimg.com/originals/e5/71/b8/e571b8e2f005a4dce30ca33dce84317e.jpg
https://animalians.wikispaces.com/file/view/TasmanianDevil.jpg/235106478/TasmanianDevil.jpg
http://www.mammalogy.org/uploads/imagecache/library_image/library/1285.jpg
The only evidence of caching I have seen in dasyurids is captive spotted tailed quolls I briefly cared for hiding rats, once in a water dish. People who extensively manage them in captivity would have much more experience. nick
One of the behavioural differences between Sarcophilus and Gulo is that the latter habitually stores food in caches, whereas such behaviour seems unrecorded in the marsupial.
It is easy to dismiss this difference along the lines that ‘because the wolverine lives in a refrigerator, obviously it is going to use the cold to preserve any surplus food’ under the snow. However, this does not stand up to scrutiny as a satisfactory explanation. This is partly because any piece of food is quite easy to preserve just by burying it. Contrary to popular assumption organic material does not quickly rot when buried; instead the mere act of burial is similar to refrigeration in retarding the process of decomposition, for the simple reason that the oxygen supply to bacteria and invertebrates is largely cut off.
Nor would it be satisfactory to claim that the main reason why Gulo is so avid in its caching of food is that it is the superior digger of the two. It is true that the fore foot of Gulo is far more powerful than that of Sarcophilus, with far longer and stouter claws. However, Gulo depends on the sharpness of its claws for climbing to escape its major enemies, which are the wolf and the brown bear. Gulo is quite unlike e.g. Mellivora because Gulo seems reluctant to dig in earth whereas Mellivora lives by digging. Gulo may look like a ‘badger’ but it is not a badger in the sense of being a habitual digger; instead if functions more like a giant marten.
In order to understand the difference between the two forms, I suggest that the most important thing to realise is that Sarcophilus is effectively the ‘top scavenger’ in its ecosystem, whereas Gulo is in competition with both the wolf and two species of bear.
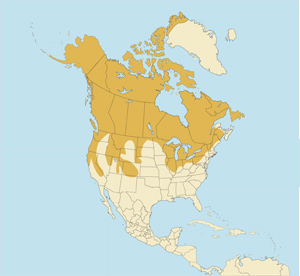
Although everyone knows that the brown bear (Ursus arctos), like the wolf (Canis lupus), ranges as far north as Alaska and Siberia, what may be more easily overlooked is that even the black bear (U. americanus) also overlaps extensively with Gulo in natural distribution. Please see the maps below.
The black bear, which weighs about 5-fold the wolverine and is omnivorous with some ability to scavenge and even to break bones in its jaws, actually occurs across most of the distribution of Gulo in North America. Admittedly bears are not major competitors for carrion or bones because they hibernate in the cold season. However, they may help to explain why Gulo habitually breaks up carcases and scatters the pieces while at the same time burying them to some degree, a pattern of behaviour of which I’ve read nothing in the case of Sarcophilus.
In the case of Sarcophilus, there is no competitor for bones and there seems never to have been.
In addition to the principle of competition for bones there is a second, possibly even more important principle involved: that in guilds of carnivorous mammals there is fighting like cats and dogs on a level that has nothing proximately to do with food. All the larger Carnivora will gladly kill each other just for the hell of it. I.e. the wolf is keen to kill the wolverine simply for existing, and even where food is not part of the immediate equation; and indeed when the wolf kills the wolverine this is usually neither because it was competing directly for food nor because it is going to eat the dead wolverine. This is hatred on a systematic scale among guild-members in Carnivora and it shapes the behaviour of the smaller members of these guilds. For example, the main reason why smaller spp. of felids have camouflage colouration is probably their fear of their larger relatives, not any need to hide from their own prey.
With all of the above in mind:
Whereas the approach of Gulo seems to be to disperse and hide any surplus bones as rapidly as possible as a individual forager, the approach of Sarcophilus seems to be to call in as many conspecific individuals as possible to share the bones as rapidly as possible.
The result is that, while food-caching is a pronounced aspect of the behaviour of Gulo, it is sociable eating which is the pronounced aspect of the behaviour of Sarcophilus. The mindset of Sarcophilus seems to be: ‘I can’t possibly finish off all the bones of this ‘roo because I’m already satiated; so let’s give them to a friend or relative so that the favour will be returned at some time in the future.’ This kind of altruism would not work well in the habitat of Gulo because calling conspecifics to the carcase would attract the competing (and downright murderous towards Gulo) wolf plus, at least in spring, summer and autumn, two species of bears.
Original distribution of Ursus americanus:
https://www.bear.org/website/images/stories/images/images-new/historic_black_bear_range.jpg
Original distribution of Gulo gulo:
http://admin.scirecordbook.org/images/species/M65.jpg
I’ve now read Owen & Pemberton (2005) carefully on the topic of Sarcophilus. I’ve found as many questions as answers, and here I’ll list some of these in no particular order. Sarcophilus is worth characterising more precisely and comparatively than anyone seems to have attempted so far.
I get the impression that Sarcophilus is in line with the semelparous smaller dasyurids, but not semelparous mainly because it is too big. It still has that kind of life history strategy but there is no precise term for its particular strategy because it does not qualify categorically as semelparous. My point is that Sarcophilus cannot be understood without considering its short lifespan and its reliance on rapid replacement of generations rather than survivorship. The proneness to cancer is interesting in the context of this limited lifespan, and fits the notion that Sarcophilus evolved essentially free of predation and limited instead by the availability of resources.
The cheek vibrissae are a special feature of Sarcophilus, in great contrast to Gulo. I’ll investigate the details further.
Sarcophilus is both smaller and less sexually dimorphic than Gulo. As a result, the largest male Gulo is five-fold the body mass of the mean female Sarcophilus, i.e. about 30 kg vs about 6 kg. Even so, I note that the largest male Gulo resembles a bear not because it is massive as much as because it has such a proportionately small head and small eyes.
Sarcophilus has pale skin (and a small patch of rather pale fur) around its eyes, which I find significant but unremarked. Whereas Gulo hides its eyes by means of a dark mask, Sarcophilus manages to advertise its eyes - despite the co-darkness of its face and its iris – to a degree not seen in related dasyurids.
If blushing of the ear pinnae occurs in social excitement, can this be seen by Sarcophilus, i.e. does it have colour vision for pink? If not, why have this physiological reaction?
The authors state repeatedly that Sarcophilus is easy to capture, but I didn’t get exactly how and why. There is a slight hint of ‘playing dead’ (as in ‘playing possum’) in Sarcophilus. Can a man likewise run down Gulo on flat ground?
If the tarsus has a bare posterior surface in Sarcophilus but not in canids, surely this is worth particular mention (see page 79).
Does Gulo ever use the fore feet to wash its face as Sarcophilus does? And does Sarcophilus uses its fore feet to pick up/handle food, particularly bare bones that it tries to crush?
Sarcophilus seems about equivalent to Gulo in digging abilities, which are modest in both and about equivalent to those of canids. In the light of this, the lack of caching of food by Sarcophilus seems significant.
If the mother in Sarcophilus really does flee with pouch-young dangling from the body and being bumped about on the ground or obstructions, shouldn’t this be stated clearly and explained? Does Sarcophilus ever carry its juveniles in its mouth, as Carnivora do? The mother in Sarcophilus carries your offspring around, after bearing them, for >15 weeks. How does the period combining gestation and the carrying of offspring in Sarcophilus compare to the gestation period in Gulo?
How do juveniles of Sarcophilus grasp the pelage of their mother? Do they use only the fore feet or the hind feet as well? Is any real dexterity involved? I get the impression that Sarcophilus uses digital grasp of the fore digits when climbing trees, rather than the traction of claws. If so, is the same true of its hind feet? Do juveniles ever ride on their mother in Gulo?
Sarcophilus seems to differ considerably from hyenas, all of which have long necks, in having ‘virtually no neck’. Why the difference?
The description of how the mouth and teeth are displayed by Sarcophilus when it reacts defensively is too vague. Is there any real fang-baring similar to that seen in the wolf or in certain felids?
It is interesting that Sarcophilus both swallows and defecates in large pieces. Is this anterior-posterior analogy a mere coincidence? I suspect that the large size of the faecal sausage is related to social marking; i.e. the animal ‘saves up’ its faeces so that it can deposit as much as possible in one go when returning to the communal latrine. Also, digestion does not seem as thorough in Sarcophilus as in hyenas. With hyenas, the faeces usually consist of paste (turning into powder) in which few if any objects can easily be discerned; with Sarcophilus, this book reports the incidence of whole recognisable paws and feet.
Is it true that Gulo is territorial despite its vast home ranges?
Underplayed in this book is that the locomotion of Sarcophilus seems specialised for flat surfaces and clear paths, and avoidance of rocks. Both contrast greatly with Gulo.
Sarcophilus presents an odd combination of inconspicuousness and noisiness. The spotted hyena, although spotted in keeping with camouflage, is not inconspicuous in this way, perhaps simply because it is so much bigger. But of which animal does Sarcophilus need to be so scared? I infer that the function of its blackness and general inconspicuousness is mainly vs its prey, i.e. for ‘hunting’. This function of colouration seems implausible in the case of hyenas as well as Gulo.
Is the gregarious eating a way of dismembering carcases by tugging, much as happens with crocodilians? It seems hard to explain any other way, because if the main function is to share food to promote the species at the level of the population, then this could be achieved by serial vocalisation AFTER satiation. This puzzle seems to have been poorly thought-out by previous authors.
Why does nobody mention whether Sarcophilus trots or not? Can Sarcophilus sit like a dog, or not?
Why does Sarcophilus soil its nest/den?
There seems no doubt that Sarcophilus is far less brainy than comparable canids, and as lacking in encephalisation as the average marsupial. What a contrast with Mellivora, which is so brainy that it is the only member of the Carnivora capable of using tools.
I intend to photo-compared the dentitions of Sarcophilus and Dasyurus, and possibly also Antechinus.
(writing in progress)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comentarios
I’m still not sure exactly how Sarcophilus climbs (whether mainly by finger-grasping or by claw-traction). However, I now have a better idea how Gulo climbs because there so many photos on the web that show this behaviour. There seems to be little doubt that Gulo climbs partly by virtue of the grappling of its relatively sharp claws on the tree surface, for both fore and hind limbs.
The claws of Gulo are by no means as sharp as the retractile claws of felids, but they are nevertheless sharp enough to grapple bark on trees. I asked questions about the dexterity of Gulo in previous emails and the first of these two photos shows Gulo ‘handling’, or at least stabilising, a large bone while eating it. I suggest that when Gulo does use its fore feet to hold food, it uses its claws rather than sensitive digit-tips to do so, thus differing from Sarcophilus.
http://wolverinepedia.zaxtor.net/WolverineClaws3.jpg
http://wolverinepedia.zaxtor.net/CLAWS_smaller.jpg
The following clearly shows this grappling in action for the fore feet.
https://thumbs.dreamstime.com/x/wolverine-tree-climbing-finnish-forest-58996353.jpg
The following shows that, under certain circumstances, the wolverine actually ‘bounds’ up the tree rather than bear-hugging its way up the tree. This suggests that the hind claws are sharp and strong enough to provide propulsive purchase on the tree trunk, something that I cannot imagine in the case of Sarcophilus. I doubt that even juvenile Sarcophilus climbs in this ‘locomotory’ way.
http://static.zoonar.com/img/www_repository1/90/34/de/10_269ea6a12ae5aea19abde3658ecb7f8b.jpg
The following again shows the hind feet in a posture suggestive of reliance on the grappling effect of sharp claws.
http://l450v.alamy.com/450v/ebx5kj/wolverine-gulo-gulo-climbing-up-a-trunk-finland-ebx5kj.jpg
The following photo is clear enough to show the tips of claws in contact with the surface of the tree.
https://s-media-cache-ak0.pinimg.com/736x/b3/7e/d0/b37ed0017baceb3f1905ea22a79ef488.jpg
In the following view, I cannot see how the uppermost forelimb could be gripping the tree trunk other than by traction of the tips of the claws.
https://s-media-cache-ak0.pinimg.com/736x/6a/0c/e2/6a0ce255f9d9b394e387eae76a77d1ea.jpg
The following view is again suggestive of grappling by means of the claws, rather than the sort of digital grasping practised by monkeys.
http://c8.alamy.com/comp/F4J82F/wolverine-climbing-a-tree-F4J82F.jpg
Here again, the distance between the animal and the tree suggests a reliance on sharp claws rather than either digital grasping or brute force in bear-hugging the trunk.
https://thumb7.shutterstock.com/display_pic_with_logo/1268695/379296619/stock-photo-wolverine-glutton-climbing-on-a-tree-379296619.jpg
Gulo is capable even of descending a tree upside down, which is surely testimony to reliance on sharp claws. Can even a juvenile Sarcophilus descend like this? I doubt it.
http://c8.alamy.com/comp/FX5XYE/wolverine-gulo-gulo-climbing-a-tree-and-is-upside-down-comming-down-FX5XYE.jpg
The following shows how Gulo can gling even to relatively smooth surfaces of trees which have lost their bark.
https://au.pinterest.com/erictrundy/wolverines/
The following again shows that, although Gulo is plantigrade, it does not apply the sole of the hind foot to the surface of the tree.
http://media.gettyimages.com/photos/wolverine-climbing-tree-side-view-picture-id200478188-001?s=170667a
The following view of the palmar surface of the manus differs from those I showed you previously, in that the fur is far shorter and thinner-looking. I assume that this is because this view was taken in summer. However, the photo once again shows how full of rubbery bumps the fore foot of Gulo really is. It is easy to imagine that these rubbery bumps provide useful friction when climbing, but hard to imagine that such broad-tipped and stiff-looking digits could perform a monkey-like grasp without the application of sharp claw-tips.
http://wolverinepedia.zaxtor.net/WolverineArm_.jpg
Here we have another clear view of the fore claws.
https://s-media-cache-ak0.pinimg.com/originals/30/28/c4/3028c4c9ec863b3562ae5761df79144e.jpg
The following photo is ambivalent about contact between claw-tips and ground in the case of the fore feet, but clearly shows how the rubbery bumps (foot-pads) keep the claws off the ground, and thus potentially sharp, in the case of the hind claws.
https://images.stv.tv/articles/w1280xh720xmFit/185928-male-woverine-at-highland-wildlife-park-supplied-by-rzss.jpg
The following does not suggest much grappling with the claw-tips, and instead suggests that on horizontal branches Gulo relies more on the friction of the rubbery pads on its feet.
https://us.123rf.com/450wm/dennisjacobsen/dennisjacobsen1504/dennisjacobsen150400041/39001990-wolverine-climbing-an-old-tree-trunk-in-its-natural-habitat.jpg?ver=6
Ditto.
http://earthjustice.org/sites/default/
After examining many photos of various spp., I’ve come to the conclusion that, although many or most mustelids share with Gulo the horizontal orientation of the pupil, Gulo is rather unusual in the length of the slot. So I don’t think the difference between Sarcophilus and Gulo can be dismissed as a case of ‘phylogenetic inertia’ in which Gulo has a horizontal-slit pupil simply because its ancestors had such a pupil. Instead, it seems that Gulo has further developed this pupil form, i.e. taken it to extremes for a small-eyed, basically nocturnal member of the Carnivora. And if so it represents not just a ‘neutral’ feature in our comparison but an example of actual evolutionary divergence w.r.t. Sarcophilus.
Gulo gulo:
http://exoticanimalz.com/wp-content/uploads/2015/03/15-wolwerine.jpg
There are a ‘standard four’ vibrissal tracts/tufts on the head of Sarcophilus. My point at the time was that Sarcophilus has more or less the same vibrissal tracts/tufts as other mammals such as foxes, but that these vibrissae are remarkably long/large. My point was not that Sarcophilus possesses any novel/unusual vibrissal tracts/tufts.
However, on closer examination of photos I have noticed an additional vibrissal tuft that I quite overlooked previously. This is located on the mid-ventral line of the lower jaw, and its long vibrissae project vertically downward. I don’t know what to call it because the short vibrissae scattered on the ‘chin’ area are already called the mandibular vibrissae. Perhaps this new tuft is already known in certain other mammals and already has a name?
The following photo clearly shows the vibrissal tuft in question, mid-way along the ventral outline of the lower jaw.
In the following photo you can see the mandibular vibrissae clearly, on the ‘chin’ of the animal. The mystacial and genal vibrissae are also clearly discernible. However, you can also see a different vibrissal tuft about mid-way along the ventral surface of the lower jaw, which I think differs from all the others in being single, not paired left/right.
The following photo clearly shows that this new vibrissal tuft is located on the mid-line of the ventral surface of the lower jaw. Its vibrissae are about as long as the supraorbital vibrissae.
In the following photo you can actually see the pale spot at which these new, mid-ventral vibrissae emerge from the pelage of the lower jaw.
The following photo shows the vibrissal tuft in question, the vibrissae of which are dark (shaded) in contrast to the well-illuminated mystacial and genal vibrissae.
The pale origin of the vibrissal tuft in question is again clearly shown in the following three photos. It is clearly non-paired and mid-ventral.
Agregar un comentario