 Favoritos de thomas_nogatz
Favoritos de thomas_nogatz





Fotos / Sonidos

Observ.
alexanderwuenscheDescripción
Sehr kleine Sippe, nur 2 cm hoch. Samen 0,4 mm lang. Ob Draba spathulata (hier nicht in Referenzliste)?
Very small clan, only 2 cm high. Seeds 0.4 mm long. Whether Draba spathulata (not in reference list here)?



Fotos / Sonidos

Qué
Chichicastle (Género Lemna)Observ.
alexanderwuenscheDescripción
@thomas_nogatz Lemna minuta, L. minor und Callitriche spec.;
Maßstab: 1 Teilstrich = 0,1 mm;
Bild 2 mit Lemna minor




Fotos / Sonidos

Qué
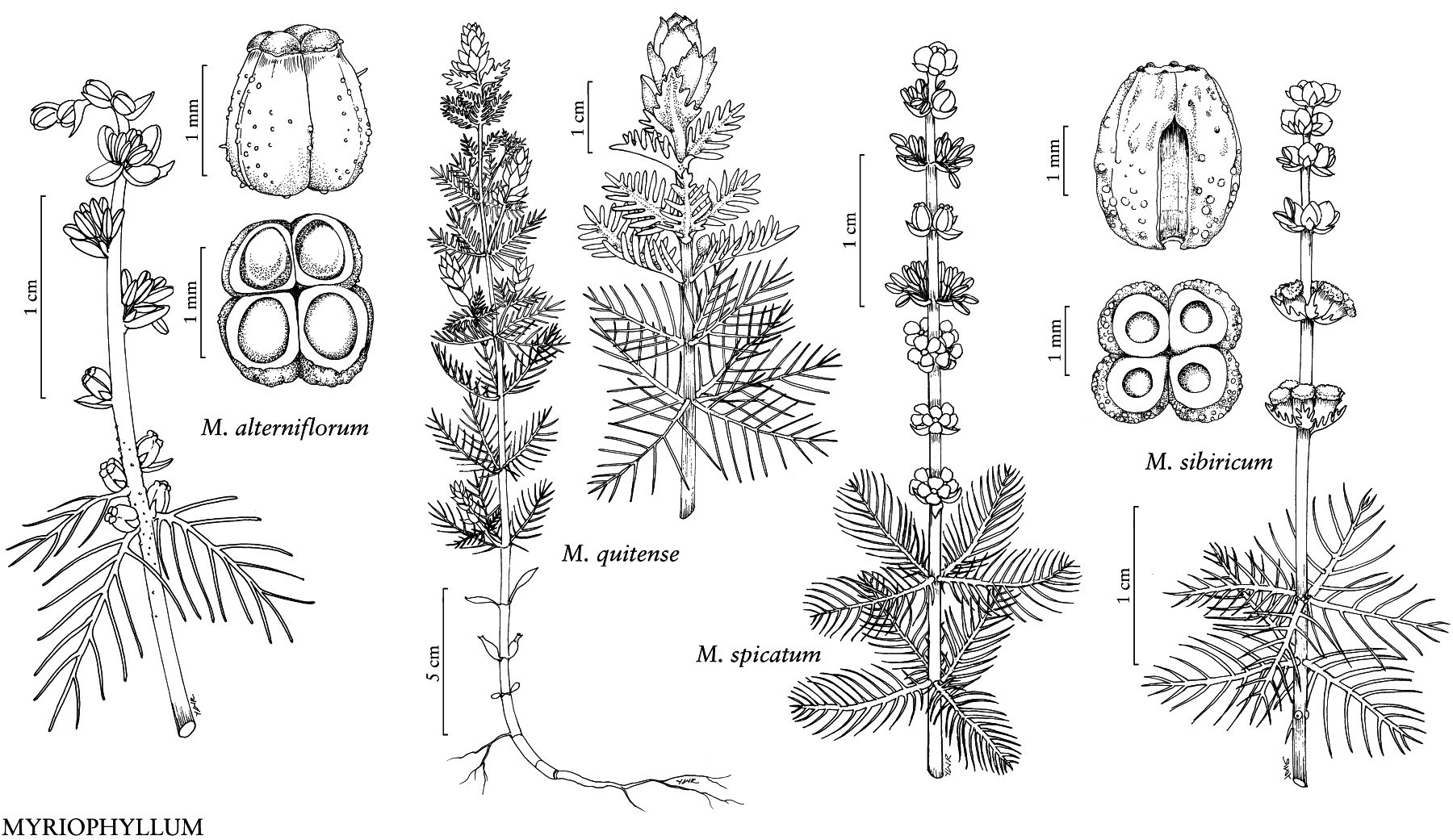
Colas de Caballo (Género Myriophyllum)Observ.
thomas_nogatzDescripción
Fundort: Dreetzsee
vgl. https://www.inaturalist.org/observations/184045196 - blühende Exemplare
und
https://www.inaturalist.org/observations/190114815 - beginnende Turionenbildung?
Merkmale:
- Anzahl Segmente pro Blatt: beidseits 6-11
- relativ weite Abstände zwischen den Blattabschnitten
- basale Blattabschnitte deutlich länger als die apikalen, oft (fast) so lang wie die Blattspindel
- Blattabschnitte zur Spitze hin gebogen
- Winkel zwischen Blattabschnitten und Mittelachse > 45°
- Blattabschnitte nicht parallel zueinander und häufig nicht in einer Ebene ausgerichtet
Merkmale lt. Literatur (s. unten) für M. sibiricum / M. spicatum:
Anzahl Segmente je Blattseite:
- 6-12 / 12-20 (Go Botany)
- 4-14 / 5-24 (Casper & Krausch 1981)
- 6-18(-24) / (20-)24-36(-42) (Scribailo & Mitchell 2022)
- 7-12 / 14-24 (Dan & Dong 2002)
- ≤14 / ≥14 (Ceska & Ceska 2012)
Winkel zwischen Blattsegmenten und Mittelachse:
- über 45° / weniger als 45° (Scribailo & Mitchell 2022)
- stumpf / spitz (Ceska & Ceska 2012)
Abstand der Blattsegmente zueinander im Blatt:
- unterschiedlich / gleichmäßig (Ceska & Ceska 2012)
Spitze der oberen Blätter:
- gerundet / abgestumpft (Go Botany)
Farbe des Stängels in trockenem Zustand:
- ± weiß bis rosa / ± rot bis hellgrün (Ceska & Ceska 2012)
Stängel unterhalb des Blütenstandes im Vergleich zur Stängelbasis:
- gleich dick / geschwollen, bis doppelt so dick (Go Botany, Casper & Krausch 1981, van de Weyer et al. 2023, Dan & Dong 2002)
- dieses Merkmal versagt bei der Untersuchung eines breiten Spektrums von nordamerikanischem Herbarmaterial (Scribailo & Mitchell 2022)
Brakteolen:
- oval, länger als breit bis so lang wie breit / nierenförmig bis fast rund, breiter als lang (Dan & Dong 2002)
Länge der Antheren:
- 1,2-1,8 mm / 1,8-2,2 mm (Dan & Dong 2002)
Bildung von Turionen = Winterknospen
- Ja / Nein (Go Botany, Casper & Krausch 1981, van de Weyer et al. 2023, Scribailo & Mitchell 2022, Ceska & Ceska 2012)
Literatur:
Native Plant Trust (2023): Go Botany. Framingham, Massachusetts
https://gobotany.nativeplanttrust.org/species/myriophyllum/sibiricum/
M. sibiricum (= M. exalbescens / M. spicatum subsp. exalbescens): mittlere Blätter mit meist 6-12 Segmenten auf jeder Seite der Spindel, obere Blätter an der Spitze abgerundet, Stängel unterhalb des Blütenstandes und nahe der Stängelbasis mit +/- ähnlichem Durchmesser, im Herbst Bildung von Turionen
M. spicatum: mittlere Blätter mit meist 12-20 Segmenten auf jeder Seite der Spindel, oberste Blätter an der Spitze abgestumpft, Stängel unterhalb des Blütenstandes dicker als an der Stängelbasis, bis zu doppelt so dick, Turionen werden nicht gebildet
Casper, S. J.; Krausch, H.-D. (1981): Pteridophyta und Anthophyta. - Ettl, H.; Gerloff, J.; Heyning, H.: Süßwasserflora von Mitteleuropa, Band 24, 2. Teil, Jena.
S. 897:
M. exalbescens: Stengel unter dem Blütenstand nicht verdickt, aufrecht. Schuppen an den Knoten des Blütenstandes 0-2, schwarz oder braun, auch im frischen Zustand undeutlich. Winterknospen (im Herbst gebildet) verkehrtlanzettlich im Längsschnitt; die schwarzgrünen Winterknospenblätter dicker als die Sommerblätter, von Oktober bis Juni vorhanden. Tragblätter der Blüten in der Regel kürzer, selten länger als die Frucht. Brakteolen 0,7-1 mm lang, eiförmig bis fast kreisrund, länger als oder so lang wie breit. Sommerblätter (10) 15-30 (40) mm lang, mit 4-14 Paaren von Blattabschnitten.
M. spicatum: Stengel unter dem Blütenstand verdickt (fast doppelt so dick wie der untere Stengel), charakteristisch gekrümmt und parallel zur Wasseroberfläche liegend. Schuppen an den Knoten des Blütenstandes 2-3, schwarz, wenigstens im frischen Zustand deutlich. Keine Winterknospen bildend, über den Winter bis auf die Wurzelkrone absterbend. Tragblätter der Blüten so lang wie oder länger als die Frucht. Brakteolen 0,5-0,8 mm lang, nierenförmig bis fast kreisrund, breiter als lang. Sommerblätter 20-40 mm lang, mit 5-24 Paaren von Blattabschnitten (gelegentlich ungeteilt).
M. exalbescens ist in Nordamerika (vielfach für M. spicatum gehalten) und auf Grönland verbreitet; in Europa bisher im nördlichen Schweden, in Finnland und in den nördlichen Teilen der Russischen SSR nachgewiesen; in Finnland und den benachbarten Teilen der UdSSR möglicherweise M. spicatum ersetzend; im Gebiet [Mitteleuropa] fehlt die Sippe.
van de Weyer, K.; Schmidt, C.; Kreimeier, B.; Wassong, D. (2023): Bestimmungsschlüssel für die aquatischen Makrophyten (Gefäßpflanzen, Armleuchteralgen und Moose) in Deutschland. Bde. 1 und 2, 4. Aufl. - Fachbeitr. LfU 119 und 120
https://offene-naturfuehrer.de/web/Myriophylliden_in_Deutschland_(Klaus_van_de_Weyer)
Bei Myriophyllum exalbescens Fernald, von dem noch keine Nachweise aus Deutschland vorliegen, ist der Stängel unterhalb des Blütenstandes nicht verdickt. Diese Art bildet Turionen aus. Myriophyllum spicatum hat einen Stängel, der unterhalb des Blütenstandes verdickt ist und bildet keine Turionen aus.
Scribailo, R. W.; Mitchell, S. A. (2022): Myriophyllum Linnaeus. - Flora of North America, Vol. 10
http://floranorthamerica.org/Myriophyllum
illustrations: http://floranorthamerica.org/w/images/e/e2/FNA10_P04_Myriophyllum_alterniflorum.jpg
M. sibiricum: Unterwasserblätter mit 6-18(-24) Blattabschnitten, diese oft unregelmäßig ausgerichtet, nicht parallel und in einer Ebene, Winkel zwischen Blattabschnitten und Mittelachse > 45°
http://floranorthamerica.org/Myriophyllum_sibiricumM. spicatum: Unterwasserblätter mit (20-)24-36(-42) Blattabschnitten, diese meist parallel zueinander und in einer Ebene angeordnet, Winkel zwischen den Blattabschnitten und der Mittelachse <45°
http://floranorthamerica.org/Myriophyllum_spicatumBis in die frühen 1900er Jahre herrschte die weit verbreitete Meinung vor, dass M. spicatum in Nordamerika heimisch und mit dem europäischen M. spicatum verwandt sei. M. L. Fernald (1919c) beschrieb M. exalbescens, um alle nordamerikanischen Exemplare von den europäischen M. spicatum zu unterscheiden, wobei der frühere Name später aufgrund des Vorrangs der Nomenklatur in M. sibiricum geändert wurde (S. G. Aiken und A. Cronquist 1988). Der erste, der das Vorkommen beider Arten in Nordamerika erkannte, war offenbar C. F. Reed (1970b). E. Hultén (1941-1950), B. C. Patten (1954) und S. A. Nichols (1975) schlugen alternativ vor, dass M. spicatum und das einheimische M. sibiricum ein Variationskontinuum bilden, was darauf hindeutet, dass es sich bei den beiden Taxa einfach um Varietäten oder Unterarten einer äußerst variablen kosmopolitischen Art handeln könnte. Auf der Grundlage einer Studie von Herbarsammlungen gingen R. Couch und E. Nelson (1985, 1992) davon aus, dass M. spicatum in den 1940er Jahren aus Europa eingeführt wurde und sich anschließend in den Vereinigten Staaten und Kanada ausbreitete. Eine neuere biogeografische Studie von cpDNA-Haplotypen deutet darauf hin, dass diese Art aus China und Korea nach Nordamerika eingeführt wurde (M. L. Moody et al. 2016).
Zu den ursprünglich von M. L. Fernald (1919c) und S. G. Aiken (1981) vorgeschlagenen und von S. G. Aiken (1981) erweiterten Merkmalen, die als zuverlässig für die Unterscheidung der beiden Arten gelten, zählen die Größe und Form der Blütenhüllblätter und Brakteolen, die Länge der Staubbeutel, die geschwollene Basis des Blütenstandes, die Farbe des Stängels in getrocknetem Material, das Ausmaß der Verzweigung und die Unterschiede bei den Teilfrüchten. Basierend auf der Untersuchung von Exemplaren für diese Arbeit, und wie von A. E. Orchard (1981) hervorgehoben, versagen die meisten dieser Merkmale jedoch bei der Untersuchung eines breiten Spektrums von nordamerikanischem Herbarmaterial. Eines der wenigen nützlichen vegetativen Merkmale zur Unterscheidung dieser Arten in den nördlichen Regionen Nordamerikas ist die Tatsache, dass M. sibiricum oft in der zweiten Hälfte der Vegetationsperiode Turionen bildet, während M. spicatum dies nicht tut (E. Hultén 1947). Das am häufigsten verwendete vegetative Merkmal zur Unterscheidung der beiden Arten ist die Anzahl der Blattsegmente der Unterwasserblätter (Fernald). Bei dem Versuch, die Pflanzen der beiden Arten zu unterscheiden, ist dies ein zuverlässiges Merkmal, aber nur, wenn die Exemplare eine niedrige (6-18) oder hohe (24-42) Segmentzahl aufweisen; die Pflanzen haben oft submerse Blätter mit einer mittleren Segmentzahl.
Molekulare Studien haben gezeigt, dass die Überschneidungen bei morphologischen Merkmalen wie der Anzahl der Blattsegmente zwischen Myriophyllum sibiricum und M. spicatum das Ergebnis einer häufigen und weit verbreiteten Hybridisierung sein können (M. L. Moody und D. H. Les 2002, 2007b; A. P. Sturtevant et al. 2009). Hybriden zwischen diesen beiden Arten können Blattsegmentzahlen von 16-28 aufweisen (Moody und Les 2007b), die sich mit den Blattsegmentzahlen von M. sibiricum (6-18) und M. spicatum (24-36) überschneiden. Eine zuverlässige Methode zur Unterscheidung dieser Taxa bei Überschneidungen in diesem Merkmalsbereich ist der DNA-Fingerabdruck (Moody und Les 2002).
Shukherdorj, B.; Shiga, T.; Batlai, O.; Wesche, K.; Ritz, C. M.; Khurelbaatar, K.; Kim, J. Y.; Jo, H. J.; Nyam-Osor, B.; Chung, G. Y.; Choi, H. J. (2019): Contribution to the knowledge on the flora of Numrug Strictly Protected Area and some parts of East Mongolia. - Journal of Asia-Pacific Biodiversity 12 (2019), 284-301
S. 288:
Abb. 4 mit Fotos vom Habitus, Turionen-Blättern, Unterwasserblättern, männlichen und weiblichen Blüten, Frucht und Samen von M. sibiricum
M. sibiricum:
7-12 Segmentpaare pro Blatt, Turionen zylindrischM. spicatum:
14-24 Segmentpaare pro Blatt
Dan, Y.; Dong, W. (2002): Taxonomic Revision of the Genus Myriophyllum (Haloragaceae) in China. - Rhodora 104/920, 396-421.
S. 398:
M. sibiricum:
7-12 Segmentpaare pro Blatt, Stängel unter dem Blütenstand fast gleich dick wie in unteren Teilen der Pflanze; Brakteolen oval, länger als breit bis so lang wie breit; Antheren 1,2-1,8 mm langM. spicatum:
14-24 Segmentpaare pro Blatt; Stängel unter dem Blütenstand fast doppelt so dick wie in unteren Teilen der Pflanze; Brakteolen nierenförmig bis fast rund, breiter als lang; Antheren 1,8-2,2 mm lang
Ceska, A.; Ceska, O. (2012): Myriophyllum sibiricum, in Jepson Flora Project (eds.) Jepson eFlora, https://ucjeps.berkeley.edu/eflora/eflora_display.php?tid=34273, accessed on October 05, 2023.
Ceska, A.; Ceska, O. (2012): Myriophyllum spicatum, in Jepson Flora Project (eds.) Jepson eFlora, https://ucjeps.berkeley.edu/eflora/eflora_display.php?tid=34276, accessed on October 05, 2023.
Myriophyllum sibiricum
Merkmale: Einhäusig; Winterknospen zylindrisch, an den Enden der nicht blühenden Zweige, meist im Herbst. Stängel: > 1 m, trocken ± weiß bis rosa. Blatt: Tauchblätter 1-3 cm, Segmente linealisch, < 20 mm, ≤ 28 pro Blatt, gekrümmt, Abstand variiert im Blatt. Blütenstand: Ähre, 3-8 cm, aufsteigend; Hüllblätter 1 - 3 mm, kürzer als die Blüten, länglich bis eiförmig, ganzrandig bis grob gezähnt.
Ökologie: Teiche, Bäche, Seen; Höhenlage: < 2600 m. Bioregionale Verbreitung: Kalifornien bis Alaska, Kanada, östliche Vereinigte Staaten, Eurasien. Blütezeit: Jun-Sep
Synonyme: Myriophyllum exalbescens Fernald; Myriophyllum spicatum L. subsp. exalbescens (Fernald) Hultén; Myriophyllum spicatum var. exalbescens (Fernald) Jeps.
Anmerkung: Erkennbar an der geringen Anzahl der Blattsegmente, dem stumpfen Winkel der Blattsegmente zur Blattrippe sowie an den zylindrischen Winterknospen.Myriophyllum spicatum
Merkmale: Einhäusig; Winterknospen 0. Stängel: > 1 m, trocken ± rot bis hellgrün. Blatt: submerse Blätter < 3 cm, Segmente ≥ 28, < 10 mm, linealisch, gekrümmt, Abstände im ganzen Blatt gleichmäßig. Blütenstand: Ähre, 4-8 cm, aufsteigend; Hüllblätter 1-3 mm, kürzer als die Blüten, lanzettlich, ganzrandig bis gezähnt.
Ökologie: Nicht häufig. Gräben, Seeufer; Höhenlage: < 2080 m. Bioregionale Verbreitung: Kalifornien bis Britisch-Kolumbien, Ostkanada, östliche Vereinigte Staaten; heimisch in Eurasien, Nordafrika. Blütezeit: Jul-Sep
Anmerkung: Erkennbar an der großen Anzahl paralleler Blattsegmente, die in spitzen Winkeln zur Spindel stehen.
.




Fotos / Sonidos

Qué
Elodea nuttalliiObserv.
piotr_kobierskiDescripción
Published:
KOBIERSKI P., RYŚ R. & PIOTROWSKI R. 2020. Materiały do flory południowo-zachodniej części województwa lubuskiego, część II. s. 82. FloraKob, Lubsko.
https://www.researchgate.net/publication/345312298
Fotos / Sonidos

Qué
Arrocillo Áspero (Leersia oryzoides)Observ.
piotr_kobierskiDescripción
Published:
KOBIERSKI P., RYŚ R. & PIOTROWSKI R. 2020. Materiały do flory południowo-zachodniej części województwa lubuskiego, część II. s. 82. FloraKob, Lubsko.
https://www.researchgate.net/publication/345312298
Fotos / Sonidos

Observ.
piotr_kobierskiDescripción
Published:
KOBIERSKI P., RYŚ R. & PIOTROWSKI R. 2020. Materiały do flory południowo-zachodniej części województwa lubuskiego, część II. s. 82. FloraKob, Lubsko.
https://www.researchgate.net/publication/345312298


Fotos / Sonidos

Qué
Najas marinaObserv.
thomas_nogatzDescripción
Merkmale, die auf Subspecies marina hinweisen:
- Blätter deutlich über 1 mm breit
- Stg bestachelt
Merkmale, die auf Subspecies intermedia deuten:
- Blattscheiden mit bis 3 Zähnchen
- Mittelnerv auf der Blattunterseite mit 2 bis 4 Stacheln
Der Tegeler See wurde 2004 in Gewässergüteklasse II eingestuft - mäßig belastet
Gefährdungen:
- Rückgang des Wasserstandes
Verbreitungskarten:
- N. marina subsp. marina: https://floraweb.de/webkarten/karte.html?taxnr=7196
- N. marina subsp. intermedia: https://floraweb.de/webkarten/karte.html?taxnr=7197
Literatur:
http://www.blumeninschwaben.de/Einkeimblaettrige/marina.htm
Buch, C.; Jagel, A.; van de Weyer, K. (2012): Najas marina L. subsp. intermedia (WOLFG. ex GORSKI)
CASPER (Hydrocharitaceae), das Mittlere Nixkraut, am Niederrhein. Erstnachweis für Nordrhein-Westfalen. Veröff. Bochumer Bot. Ver. 4(4): 38-43. https://www.botanik-bochum.de/publ/OVBBV4_4_BuchJagelvandeWeyer_Najas_intermedia.pdf
Berliner Senatsverwaltung für Stadtentwicklung (2004): 02.03 Biologische Gewässergüte (Trophie): https://www.berlin.de/umweltatlas/_assets/wasser/gewaesserguete-trophie/de-texte/k203.pdf
Rüegg, S. (2000): Genetic and phenotypic differentiation of Najas marina L. s.l. in relation to environmental conditions. Diss. TU München: https://mediatum.ub.tum.de/doc/1579930/1579930.pdf
Rüegg, S.; Braeuchler, C.; Geist, J.; Heubl, G.; Melzer, A.; Raeder, U. (2019): Phenotypic variation disguises genetic differences among Najas major and N. marina, and their hybrids. In: Aquatic Botany, Vol. 153: S. 15-23: https://www.sciencedirect.com/science/article/abs/pii/S0304377018301281














Fotos / Sonidos

Qué
Geranium aequaleObserv.
epsilonDescripción
Teilfrüchte glatt, im Gegensatz zu G. molle
Blütenblätter an der Spitze ausgerandet, im Gegensatz zu G. rotundifolium
Stängel mit langen Haaren, im Gegensatz zu G. pusillum
Blütendurchmesser ca. 1 cm, im Gegensatz zu G. pyrenaicum ( ca. 1,5 cm)
Stängelblätter wechselständig, im Gegensatz zu G. pyrenaicum
http://www.blumeninschwaben.de/Zweikeimblaettrige/Storchschnabelgewaechse/ohne_granne.htm#6



{kind=link}